Image from Bio Rad
As a follow up of Thursday's lab, we stained the gels with Coomasie G-250 stain and then air dried them for analysis.

(click on pic for a full size image)
-----------------------


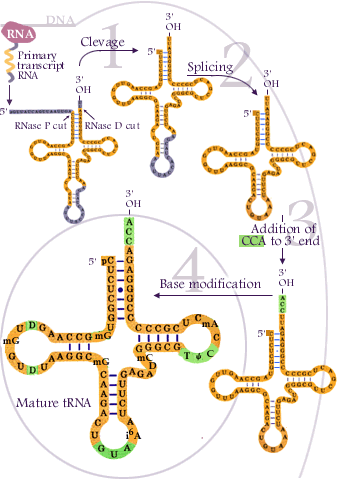
Today we finished chapter 11 on RNA processing. We focused mainly on alternative splicing and how it produces different mRNA molecules by transcribing the same gene.
We also discussed processes like base modification, base substitution, RNA editing, and RNA degradation.
After finishing the chapter we discussed students' impressions on Dr. Reijo-Pera's Keiser lecture yesterday evening and expectations for her talk in our class tomorrow...!!! (expectations from the talk itself and about students' interaction with Dr. Reijo-Pera)
Tomorrow:
Dr. Renee Reijo-Pera, from Stanford University, will give a lecture on human preimplantation development and gene expression and pathways during the first few days of development. We will be joined by students in Dr. Aulthouse's Developmental Anatomy class, and potentially Dr. Walden's CLS program so the room will be packed. The talk will be as exciting as the Keiser lecture and having two-three classes in the audience will make the discussion more interesting and lively...!
Students should be ready to ask questions to, and engage in a discussion with, Dr. Reijo-Pera. Please check the following links:
It will be an exciting day. Take advantage of it!